B-клетки – основа гуморальной части адаптивного иммунитета. Я хочу продолжить тему базовой информации об иммунной системе, а в конце расскажу про небольшую революцию в понимании работы B-клеток. Предыдущую заметку рекомендую почитать, так как текст этой подразумевает знание предыдущего материала. В будущем я буду довольно глубоко зарываться в иммунологию и генетику. Хочу, чтобы на моем сайте была минимальная база, на которую я всегда смогу дать ссылку любопытному читателю. Поэтому помимо привычных статей у меня будут такие общеобразовательные вещи.
Примерный план:
- B-клетки и функции антител;
- Строение и классы антител;
- Активация B-клеток CD4+ клетками;
- Реакции герминативных центров;
- Относительно недавние открытие в области гуморального иммунитета;
B-клетки и антитела
B-клетки – это клетки адаптивной иммунной системы. Их основная особенность – способность секретировать антитела.
Антиген – всё, что угодно. Любая молекула, которая может связаться с антителами.
Антитело – белковая молекула, выполняющая ряд важных функций. Особенность антител в том, что их может быть 10˄10 – 10˄11 различных разновидностей. Антитела могут связываться практическим с любым антигеном. В отличие, допустим, от T-клеток, для активации которых нужны пептиды, презентованные на MHC-белках;
Функции антител:
Нейтрализация. Антитело может связаться с патогенным компонентом (допустим, вирусом), не давая ему связываться с клеточными целями;
Опсонизация. Фагоциты могут не видеть часть патогенов. В том числе антитела обволакивают антигены, делая их доступными для фагоцитоза;
Фиксация комплиментов. Комплименты в свою очередь способствуют воспалению, опсонизируют патогены или с помощью C9 MAC (membrane attack complex) разрушают клеточную стенку бактерий (лизис), тем самым их уничтожая.
Цитотоксичность, опосредованная антителами. NK-клетки и цитотоксичные Т-клетки могут присоединяться к антителам, выпуская смертоносные гранулы в зараженные (в идеале) клетки.
Строение антитела
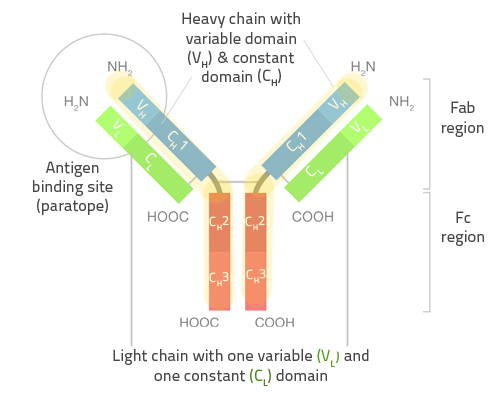
Антитело состоит из двух Heavy Chains и двух Light Chains.
Fc часть неизменяема, с ней связываются другие клетки, комплименты итд.
Fab часть содержит у N-терминалов гипервариативные регионы. V(D)J рекомбинация гена, контролирующего 3 петли (loops) у каждого из гипервариабельных регионов обеспечивает все многообразие антител.
Антигены связываются с Antigen binding site.
Углубляться не будем, чтобы не забыть про основную тему.
Классы антител (иммуноглобулинов)
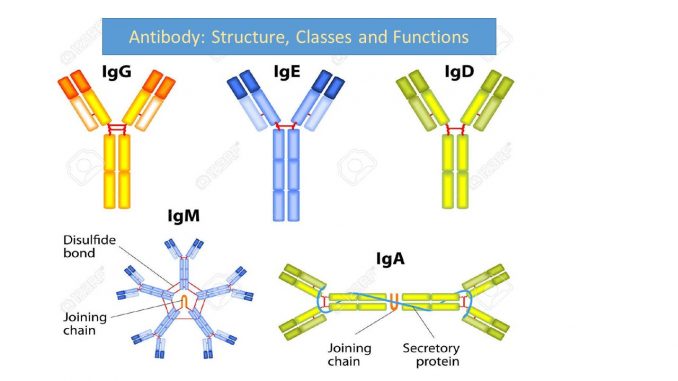
IgM – это базовый иммуноглобулин. Визуально его отличает J-chain, соединяющая 5 антител в одно. Изначально B-клетки секретируют именно IgM и только последующие события позволяют менять этот изотип на другой.
IgG – рабочая лошадка антител, самый распространенный иммуноглобулин. Обладает большинством функций, которые мы ассоциируем с антителами. В том числе наследование антител от матери детям.
IgA – также обладает J-chain, в данном случае устойчивой к протеазам. Что определяет место действия этого Ig – желудочно-кишечный тракт.
IgE – активирует эозинофилы, важно для реакции на паразитов, также участвует в аллергических проблемах.
IgD – менее важный и менее понимаемый Ig. Играет роль в том числе в работе легких.
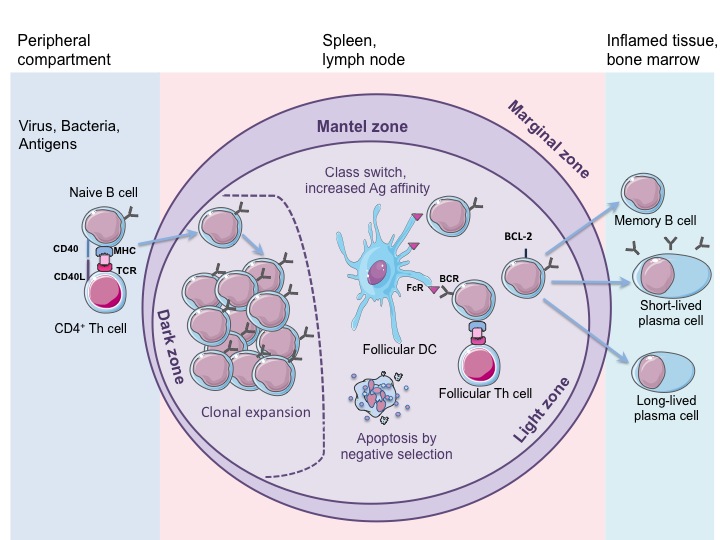
B-клетки. Активация CD4+ клеткок в лимфоузлах
B-клетки и T-клетки происходят из hematopoietic stem cell (гематопоэтические стволовые клетки или гемоцитобласты). Но их судьба разделяется после common lymphoid progenitor, общего предка всех лимфоцитов. B-клетки большую часть «взросления» проходят в костном мозге, а Т-клетки в тимусе. Возмужавшие, но все еще незрелые (immature) клетки обоих типов затем отправляются во вторичные лимфоидные органы, в первую очередь в лимфатические узлы.
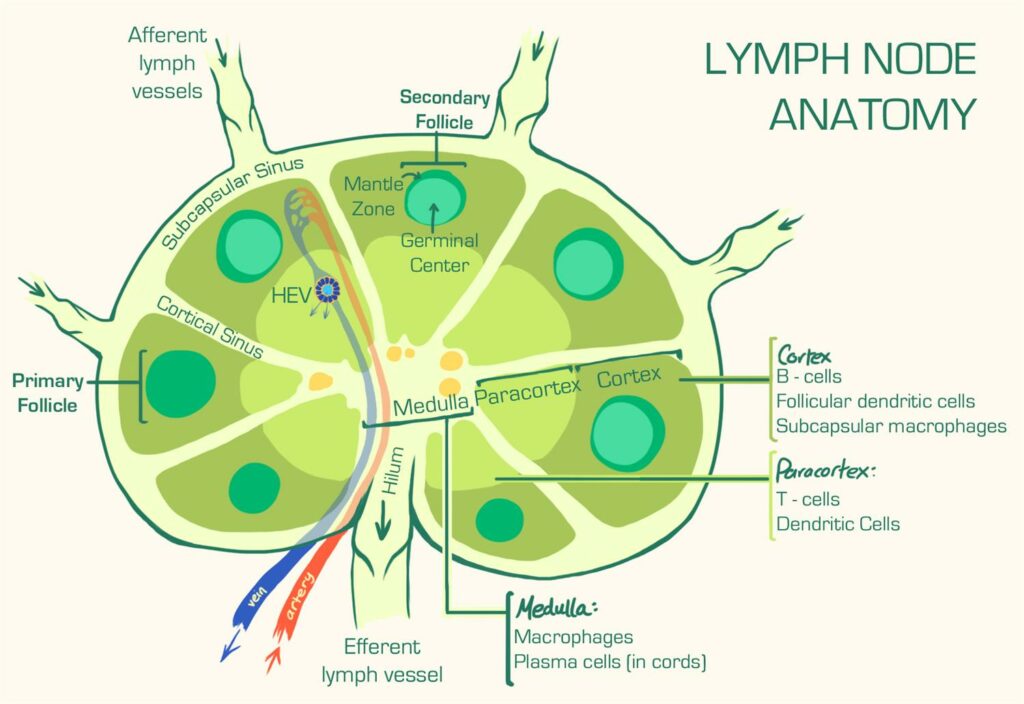
B-клетки населяют фолликулярную часть лимфоузлов, в том время как Т-клетки населяют межфолликулярное пространство, где (как помните) активируются дендритными клетками.
Сближают клетки хемокины Т-клеток и CCR7 рецепторы B-клеток, знакомый нам по активации дендритных клеток.
B-клетки также являются антиген-презентующими клетками (вместе с DC и МФ). Они интернализируют патоген, расщепляют его белки на пептиды в лизосомах, затем представляют пептидные фрагменты патогена на MHC II белках. Раз речь идет об MHC II, то и активировать B-клетки будут CD4+ (T helper) лимфоциты.
Пока у нас B-клетки, презентующими пептид на MHC II белках и с активированным CCR7 рецептором, позволяющим воспринимать хемокины Т-клеток и мигрировать в межфоликулярное пространство.
Затем нам нужна Т-клетка с рецептором (TCR, T cell receptor), аффинитивным представленному пептиду.
В данном случае также работает принцип «двойного рукопожатия» и мембранный лиганд CD40L Т-клеток должен соединиться с CD40 белком B-клеток.
После этого обе клетки мигрируют в герминативные центры (germinal center), где произойдет реакция герминативных центров (germinal center reaction).
Germinal center reactions
Активированная Т-клетка (назовем ее для простаты фолликулярная Т-клетка, fTh) от обычной будет отличаться:
- Белком CD40L;
- CXCR5 (он же BLR1, он же CD185)
CXCR5 рецептор позволяет B-клетке и Т-клетке вместе мигрировать в герминативный центр, где и происходит самое интересное.
Действующие лица реакции герминативных центров:
- B-клетки;
- Фолликулярные Т-клетки;
- Фолликулярные дендритные клетки.
B-клетки:
- Проходят реакцию массивную клональную экспансию;
- Которая сопровождается соматической гипермутацией (в 10^5 – 10^6 бодрее, чем мутации ДНК) частей ДНК, отвечающих за антигены;
- Соматическая гипермутация приводит к affinity maturation, резкому росту аффинитивности антигенов патогену;
- Антитела за счет генной рекомбинации (разрезке и склейке) проходят isotype switching, антитела/иммуноглобулины меняют базовый изотип IgM на какой-то другой из вышеописанных.
Фолликулярные T-клетки:
- CD40L как раз и позволяет B-клеткам инициировать процедуру смены изотипа иммуноглобулина;
- Также проходят клональную экспансию, но в значительно меньше количестве, чуть дальше вы поймете, почему
Фолликулярные дендритные клетки:
Презентуют антиген B-клеткам; Во время клональной экспансии и соматической гипермутации B-клеток аффинитивность патогену будет меняться не только в большую, но и в меньшую сторону. Фолликулярные дендритные клетки с помощьюх химических сигналов добиваются того, что в живых остаются только B-клетки с рецепторами/антителами высоко-аффинитивными патогену. B-клетки с низкой и нормальной аффинитивностью погибают. Так и происходит affinity maturation.
Результаты germinal center reaction:
- Долгоживующие плазма-клетки (long-lived plasma cells), секретирующие антитела, высоко аффинитивные патогену;
- Memory B-cells, клетки гуморальной памяти; они неактивные, но при контакте с патогеном «оживают» и начинают секретировать антитела;
- Isotype switching (смену изотипа иммуноглобулина) c IgM на IgG и другие типы.
Под конец позволю себе немного лирики про разнообразие B-клеток в герминативных центрах
Иногда в науке бывают лже-аксиомы. Какие-то вещи принимаются по умолчанию, хотя никогда толком не обсуждались и не доказывались. Всегда считалось, что в germinal center reaction участвует 1 (ну изредка 2) разных клона B-клеток (в смысле презентующих разные пептидные части патогена).
Visualizing Antibody Affinity Maturation in Germinal Centers – в этой по-настоящему крутой статье авторы задались подобным вопросом, используя интересные технологии клеточной визуализации и смогли продемонстрировать однозначный результат, опровергающий лже-аксиому.
Если вы внимательно посмотрите картинки, а еще лучше если почитаете текст, то увидите, что в герминативном центре десятки разные клонов B-клеток. Это говорит о том, что наша иммунная система нападает не просто на какую-то отдельную «пептидную сигнатуру», а атакует вражеские белки «с разных сторон».
B-клетки и итоги
Так как выше скорее экспресс-вводная по функции B-клеток и реакции герминативных центров, то уместны не выводы, а напоминание того, что нужно знать применительно к гуморальному иммунитету и этой заметке:
- Ключевая функция B-клетки;
- Что такое антигены;
- Что такое антитела;
- Функции антител:
- Опсонизация;
- Нейтрализация;
- Фиксация комплиментов;
- Цитотоксичность, опосредованная антителами;
- Структуру антитела:
- Fab / Fc; что связывается с антигенами, с чем связываются другие иммунные клетки;
- Heavy chains, Light Chains;
- Antigen binding site;
- Гипервариативный домен антител; V(D)J рекомбинация;
- Классы антител: IgG, IgM, IgA, IgE, IgD и их основные особенности;
- В самом примитивном виде структуру лимфатического узла (где какие клетки);
- Антиген-презентующая функция B-клеток;
- CCR7 и миграция в межфолликулярное пространство;
- Активация Т-клеток (MHCII + TCR; CD40 + CD40L);
- Отличия активированной (фолликулярной) Т-клетки от обычной:
- CD40L;
- CXCR5;
- За счет чего (CXCR5) клетки потом мигируют в герминативный центр;
- Клетки герминативных центров и их функции;
- Основные процессы внутри герминативных центров (соматическая гипермутация, affinity maturation, isotype switching);
- Итоги реакции герминативных центров (плазма-клетки, memory B cells, isotype switching);
- Количество видов B-клеток в герминативных центрах: устоявшееся мнение и последние исследования.